
Die genetische Entwicklung der Europäischen Eibe (Taxus baccata) während des Quartären Eiszeitalters
Eine Zusammenfassung von Mayol, et al. 2015, „Adapting through glacial cycles: insights from a long-lived tree (Taxus baccata)“
Fred Hageneder, Mai 2015
2015 erschien im New Phytologist eine Arbeit, in der 14 Wissenschaftler aus Spanien, Frankreich, Italien und Slowakien die Ergebnisse ihrer Studien zur genetischen Entwicklung der Europäischen Eibe (Taxus baccata) durch die glazialen Zyklen des Quartärs vorstellten.
Die Untersuchung fand Belege, dass die Eibe Europa von Osten her besiedelt hat und dass die Art sich zu Beginn der quatärneren Vereisungen vor ca. 2,2 Mio. Jahren in zwei Gruppen (westliche, östliche) teilte. Die Untersuchung entdeckte außerdem ein erhebliches Maß an umweltbedingter Anpassung am Ursprung der genetischen Abzweigung während der Interglaziale.
Da diese Vorgänge auch in anderen Arten üblich sind, bietet diese Studie neue Untersuchungsmethoden, um die Auswirkungen der quatärneren Klimafaktoren auf die heutigen Verbreitungsmuster der genetischen Vielfalt zu erforschen.
Die Vereisungen („Eiszeiten“) und die dazwischen liegenden Warmzeiten (Interglaziale) des Quartärs (dem gegenwärtigen und jüngsten geologischen Zeitabschnitt, der vor 2,58 Mio. Jahren begann) sind seit langem bekannt als der Haupteinfluss auf die geographische Verteilung europäischer Arten und auf die Muster ihrer genetischen Unterschiede. Aber ihr Beitrag zur geographischen Isolation und/oder umweltbedingter Anpassung durch die Erzeugung genetischer Abweichung in Populationen wurde bisher nicht näher untersucht. Diese Studie benutzte einen langlebigen Baum als eine Schablone, um die Wirkung quatärnerer Klimaänderungen auf die Genvielfalt über neutrale („Isolation durch Entfernung“, IBD) und selektive („Isolation durch Adaption“, IBA) Prozesse zu untersuchen. Bäume sind durch ihre Langlebigkeit als Modellarten besonders geeignet, weil…
• sie über riesige Flächen mit einem weiten Spektrum an biotischen (Pflanzengemeinschaft, Äsungsdruck, u.a.) und abiotischen (Terrain, Boden, Position, Niederschlag usw.) Faktoren verteilt sind,
• sie örtliche Anpassung an umweltbedingte Gradienten aufweisen,
• die Merkmale ihrer Lebensgeschichten (z.B. Langlebigkeit, überlappende Generationen, ausgedehnte jugendliche Phase) die Effekte von Veränderungen der Genstruktur abpuffern.
Andere Untersuchungen (an Eichen, Q. engelmannii, Q. lobata) haben bereits gezeigt, dass „das genetische Signal vergangener Klimaperioden in Organismen mit großer effektiver Populationszahl und langen Generationsspannen über große Zeiträume fortdauern kann.“
Materialien and Methoden
Um den wahrscheinlichsten Zeitverlauf der Erscheinung umweltbedingter Barrieren für den Genfluss (und daher auch die Rolle ihres tatsächlichen Beitrags zur „Isolation durch Adaption“, IBA) zu ermitteln, wurden verschiedene paläoklimatische Datenbanken benutzt und einzeln getestet. Die Methoden der jüngst entwickelten Annähernden Bayesischen Computation (ABC, approximate Bayesian computation) halfen, komplexe demographische Szenarien zu durchleuchten und ihre Zeitabläufe zu schätzen. Schließlich wurden die zugänglichen Klimainformationen von drei Zeitabschnitten –
• das letzte Interglazial (LIG, ca. 120.000–140.000 BP*),
• das letzte glaziale Maximum (LGM, ca. 21.000 BP) und
• heutige Bedingungen (PRE, ca. 1950–2000) –
* BP = before present, vor der Gegenwart
benutzt, um die relative Bedeutung gegenwärtiger und vergangener Klimabedingungen auf die beobachteten Muster genetischer Variation zu bewerten. Die Verbindung dieser Ansätze half zu bestimmen, ob die stehenden Muster genetischer Abweichung das Ergebnis von geschichtlicher Isolation sind oder von lokaler Anpassung an ökologisch unterschiedliche Gebiete.
Sampling, DNA-Extraktion und nuclear microsatellite genotyping: Insgesamt 4992 Untersuchungsproben (n = 1–60 pro Lokalität, Durchschnitt 21) wurden an 238 Standorten gesammelt, die das gesamte Verbreitungsgebiet von Taxus baccata L. abdecken. Reine DNA wurde aus 50 bis 100 mg trockenen Laubes isoliert. Sieben Primer-Paare, speziell für Taxus baccata zur Amplifikation von nuclear microsatellites (nuSSRs) entwickelt, wurden zur Gen-Analyse eingesetzt.
Chloroplast DNA sequencing: Sechs Chloroplasten-Regionen plus die trnS-trnQ spacer-Region wurden getestet, aber nur drei Regionen konnten erfolgreich amplifiziert werden: rbcL, trnS-trnQ und trnLtrnF. Die amplifizierten Produkte wurden auf Polymorphismus geprüft (screened) und gereinigt, und ihr Erbgut wurde von beiden Enden her entschlüsselt. Sequenzen aus früheren Studien wurden von Gen-Bank gedownloaded und mit den neu gewonnenen Sequenzen ausgerichtet.
Genetische Diversität: In allen Populationen mit mindestens acht Individuen (195 Standorte, n = 4829) wurden die beobachtete und die erwartete Heterozygotie als auch der Reichtum an Allelen* berechnet und das Koppelungsungleichgewicht unter allen Genlocus-Paaren in jeder Population bewertet.
* Die meisten zellulären Organismen haben zwei Sets von Chromosomen, d.h. sie sind diploid. Diese Chromosomen nennt man homogene Chromosomen. Wenn beide Allelen an einem Genlocus (oder Gen) des homogenen Chromosoms gleich sind, nennt man den Organismus homozygot bezüglich dieses Gens, sind sie verschieden, spricht man von Heterozygotie. (nach Wikipedia engl.)
** Als Allele (von gr. allélon „einander, gegenseitig“) bezeichnet man die alternativen Formen eines Gens.
Demographische Geschichte: Das ABC-Rahmenwerk wurde auf die nuSSRs-Daten angewandt, um die demographische Geschichte von Taxus baccata abzuleiten. Ein „klares westwärts gerichtetes Gefälle sich vermindernder Vielfältigkeit von Iran zum Mittelmeerraum wurde … festgestellt, welches auf ein Besiedelungsmuster hinweisen könnte“, das von Ost nach West verlief. Darum wurden vier Szenarien durchgespielt, die alternative Hypothesen zu zwei bzw. drei Genpools berücksichtigten:
A) ein „sekundärer Kontakt“ zu einem späteren Zeitpunkt, an dem sich der westliche und der östliche Genpool noch einmal mischten (Admixed = Gemischt),
B) eine klar westwärts gerichtete Entwicklung („Besiedelung“), bei der Admixed lediglich aus dem westlichen Genpool hervorging,
C) ein „sekundärer Kontakt“ (wie bei a), aber mit der Abzweigung des östlichen Pools von einem noch älteren in Iran,
D) (B und C kombiniert, d.h.) drei Pools (westlich, östlich, iranisch), und Admixed als Abzweigung aus dem westlichen Genpool.
Mit anderen Worten: Die Szenarien A und B untersuchten zwei genetische Pools, C und D drei. Während A und C einen sekundären Kontakt des westlichen und östlichen Pools (Admixed) testeten, untersuchten B und D die Möglichkeit, dass Admixed sich lediglich aus dem westlichen entwickelt haben könnte.
Eine Population galt als Admixed, wenn die Proportion der Individuen, die zur westlichen oder östlichen Gruppe gehörten, kleiner als 70 % war.
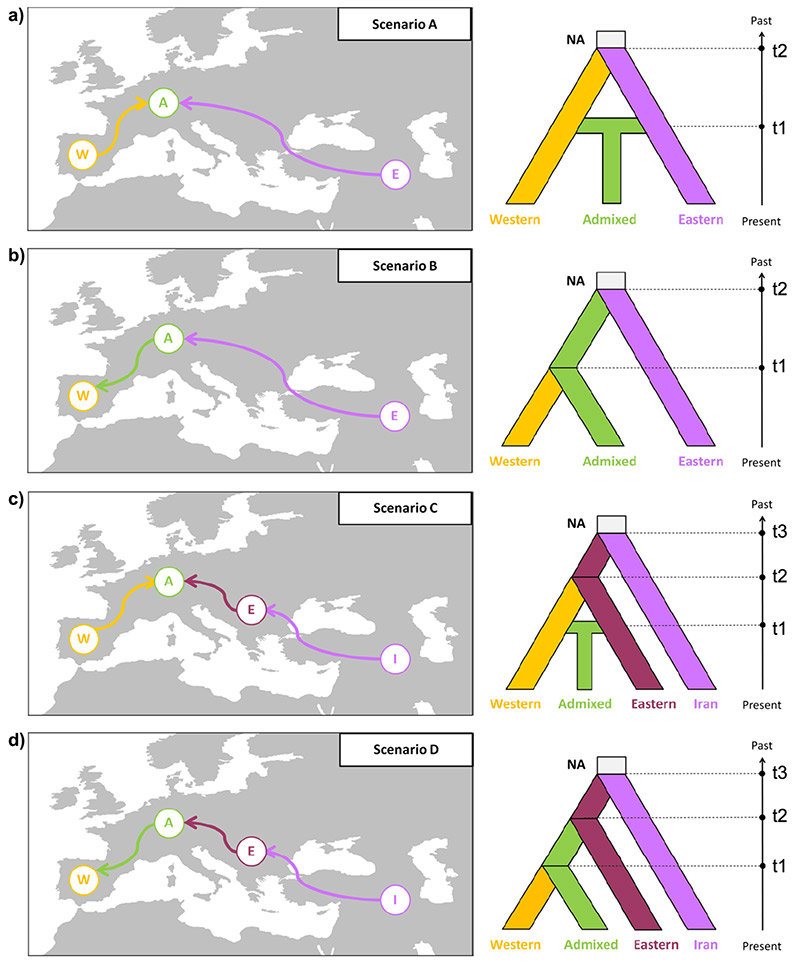
NA = Ahnen, effektive Populationsgröße; t1–t3 = Zeitpunkte der Abweichung
Da das Zusammenbringen von Daten aus unterschiedlichen Populationen die Rückschlüsse auf die demographischen Parameter verfälschen kann, wurden zehn verschiedene Populations-Sets konstruiert, von denen jedes ca. 500 Individuen enthält, die ca. 10 % des gesamten Datenbestandes repräsentieren. Jedes Set bestand aus ca. 200 Individuen des östlichen Pools, ca. 200 Individuen des westlichen Pools und ca. 100, die zu Admixed gehören – diese wurden die „500-Proben Datensets“ genannt. Für die Szenarien C und D wurden zwei weitere Datensets mit vier Populationen konstruiert, bei denen der Iran-Pool (Iran, Georgien) unabhängig vom östlichen Genpool ist. Diese Datensets bestanden aus jeweils ca. 200 Individuen der westlichen, der östlichen und der iranischen Gruppe sowie ca. 100, die zu Admixed gehören – diese wurden die „700-Proben Datensets“ genannt.
Eine Million Simulationen wurden für jedes Datenset gerechnet.
Umweltbedingte Faktoren: Klima-Informationen der WorldClim Datenbank wurden angewandt auf die Standorte der 195 Populationen, um die Wirkung vergangener und gegenwärtiger Klimabedingungen auf die gegenwärtige genetische Struktur zu bewerten. Dies wurde parallel durchgeführt mit zwei Modellen: dem Community Climate System Model (CCSM) und dem Model for Interdisciplinary Research on Climate (MIROC).
Ergebnisse
Die Wechselbeziehung zwischen genetischer und geographischer Distanz war hochsignifikant, was auf die Existenz eines IBD-Musters (isolation-by-distance = Isolation durch Entfernung) hinweist. Die genetischen Distanzen vergrößerten sich nach Westen hin, was eine größere Nähe der östlichen Populationen zur hypothetischen „Ur-Population“ nahelegt.
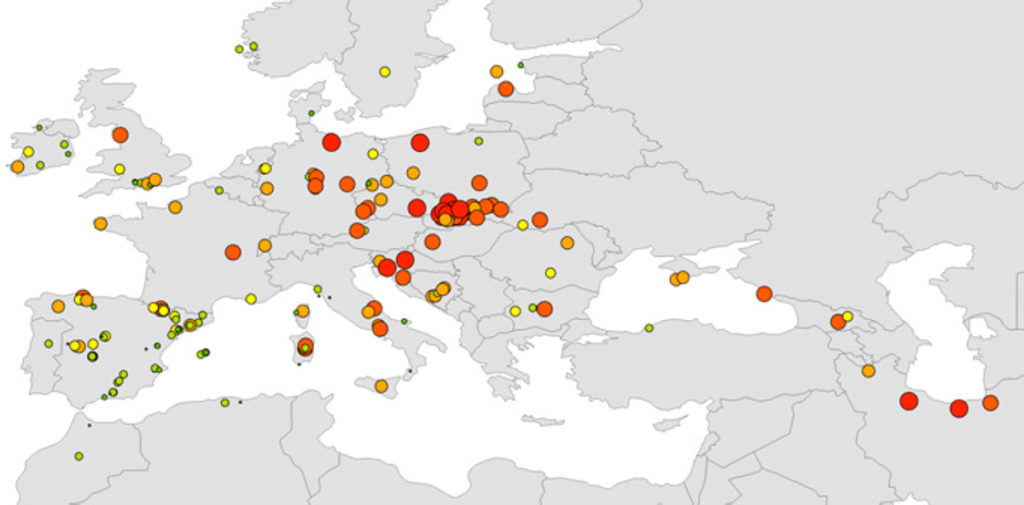
AR = Allelenreichtum; HE = unverfäschte erwartete Heterozygotie. Die Werte für AR und HE werden angedeutet durch Größe der Kreise bzw. Farbabstufung (rot > orange > gelb > grün).
Das wahrscheinlichste Szenario für die „500-Proben Datensets“ war Szenario A, „mit starkem Rückhalt in fast allen Fällen“. Entsprechend bestätigten die Simulationen mit den „700-Proben Datensets“ eindeutig das Szenario C. Weitere Testlaufprozeduren brkräftigten die Zuverlässigkeit von Szenario C als das wahrscheinlichste von allen.
Nach diesem Modell (Szenario C) müsste sich die Trennung zwischen den iranischen und den europäischen Proben etwa vor 6 Mio. Jahren* ereignet haben (bei einer angenommenen Generationsspanne von ca. 100 Jahren für Taxus baccata). Die nachfolgende Auseinanderentwicklung der westlichen und östlichen Gruppen müsste vor 2,2 Mio. Jahren** stattgefunden haben, und die spätere Vermischung vor ca. 200.000 Jahren.***
* 90 % glaubhafte Intervalle: 1,35 bis 14,78 Mio. Jahre BP
** 90 % glaubhafte Intervalle: 0,5 bis 7,5 Mio. Jahre BP
*** 90 % glaubhafte Intervalle: 50.000 bis 800.000 Mio. Jahre BP
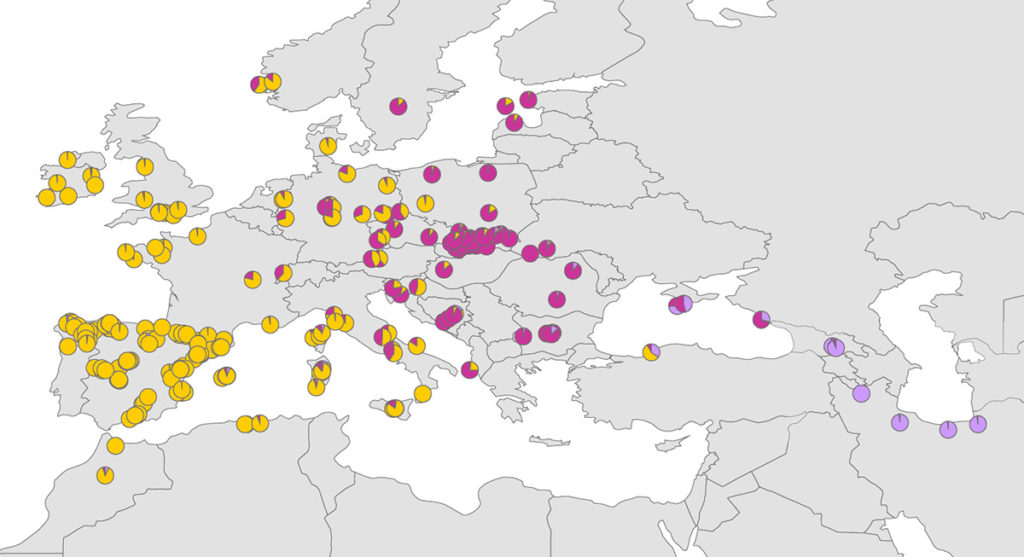
Die Tortendiagramme zeigen die ungefähren Proportionen der Zugehörigkeit zu den genetischen Gruppen, die hier in verschiedenen Farben dargestellt sind.
Die klimatische Untersuchung zeigte, dass im Durchschnitt die Populationen des westlichen Pools geringere jahreszeitliche Temperaturunterschiede, wärmere Temperaturen (Jahresdurchschnitt, -minimum und -maximum), höhere Temperaturen in der trockensten Jahreszeit sowie weniger Niederschlag während der wärmsten Jahreszeit erfuhren. Es gibt außerdem für alle berechneten Zeitabschnitte wenig Überlappung ihrer vorausgesagten Verbreitung, besonders während der beiden Interglaziale, was darauf hinweist, dass die westliche und die östliche Gruppen seit sehr langer Zeit ökologisch unterschiedliche Regionen bewohnt haben.
Diskussion
Die nuclear microsatellites im Genpool von Taxus „scheinen den Abdruck uralter Zeiten erhalten zu haben“. Des weiteren „wurde eine fast vollständige Abwesenheit von Variation und von räumlicher Struktur in den Chloroplasten-DNA-Markern beobachtet, in Übereinstimmung mit dem langsamen Nukleotidaustausch in Chloroplasten, wie er in Koniferen bekannt ist“.
Die einstige Wanderung aus dem Osten wird außerdem belegt durch die nach Westen hin abnehmende genetische Vielfalt sowie die nach Westen hin zunehmenden FST-Werte (genetische Distanz). „Im Einklang mit unseren Ergebnissen setzen neuere Studien die Entstehung von Taxus in Nordamerika oder Südwestchina während der späten Kreidezeit bis zum Mittleren Eozän (66,55 bis 11,22 Mio. Jahre BP) an, von wo aus sich die Gattung in die heutigen Gebiete verbreitete. Sie erreichte Europa wahrscheinlich über die iranisch-turkanische Region, welche als Schlüsselregion für die [allgemeine] Besiedelung des Mittelmeerraumes bezeichnet wurde (…). Dies geschah vermutlich vor dem Unteren Miozän, wie durch die älteste Fossilienfunde (16–23 Mio. Jahre BP) angedeutet wird“.
Die ABC-Simulationen legen die Trennung des europäischen vom iranischen Genpool auf ca. 6 Mio. Jahre BP (obwohl die zu 90 % glaubhaften Intervalle groß sind: durchschnittl. 1,35 bis 14,78 Mio. Jahre BP). Verringerter Genfluss ist für Taxus eine naheliegende Annahme und wird durch hohe Werte für paarweise genetische Differenzierung in dieser Studie unterstützt.
„Die einstige Trennung wird auch ersichtlich durch die Ergebnisse der Chloroplasten-DNA-Marker, weil die einzigen ausgeprägten Haplotypen* am äußersten östlichen Ende (Iran) des Verbreitungsgebietes gefunden wurden. Das deutet an, dass beide Gruppen vor langer Zeit isoliert wurden. Es wird außerdem bestätigt durch die signifikant hohe Anzahl privater Allelen in den nuclear microsatellites im östlichen Pool, besonders in den Populationen in Iran und Georgien.“
* Ein Haplotyp (Abkürzung für haploider Genotyp) ist eine Ansammlung spezifischer Allelen (d.h. spezifischer DNA-Sequenzen) in einem Cluster eng verbundener Gene auf einem Chromosomen, die mit hoher Wahrscheinlichkeit zusammen vererbt werden. D.h. sie erhalten sich wahrscheinlich als Sequenz über viele Generationen hinweg“ (nach Wikipedia engl.)
Diese Arbeit bekräftigt die Hypothese, das sowohl die Geographie als auch das Klima – besonders in den Warmzeiten – eine bedeutsame Rolle in der Formung der genetischen Struktur von Taxus in Europa innehatten. „Die Abweichung zwischen den westlichen und den östlichen Gruppen kann erklärt werden durch das Verharren in räumlich isolierten Refugien während der Vereisungen. Allerdings zeigten Artverbreitungsmodelle (Abb. 4) fast keine Überlappung beider Gruppen im Hinblick auf ausgeprägte klimatische Bedingungen, insbesondere während der Interglaziale (…).“
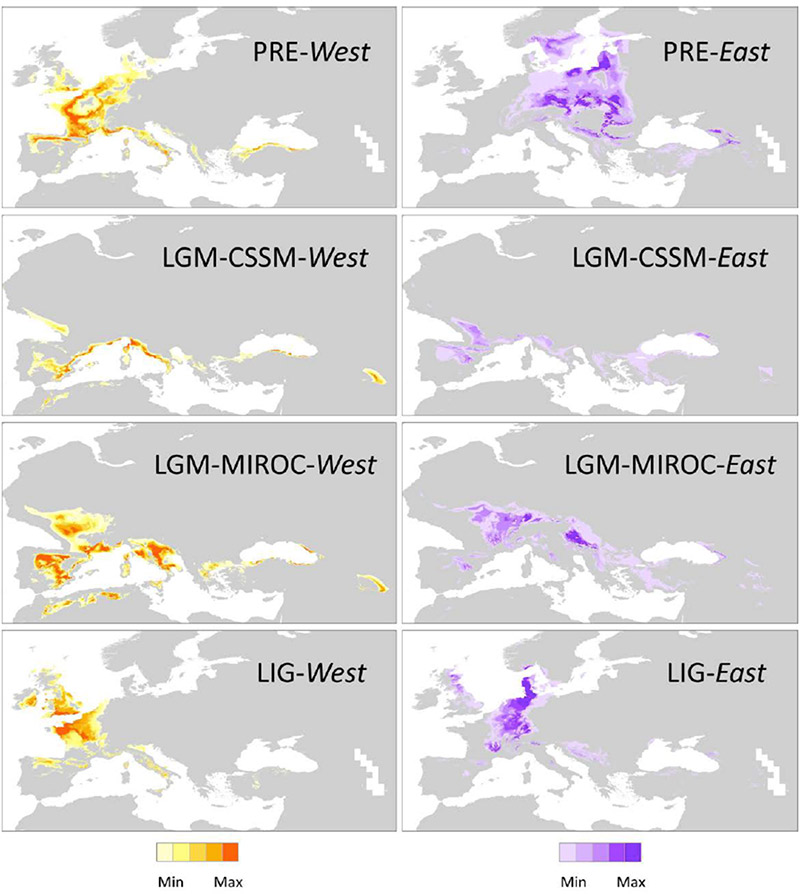
LIG, letztes Interglazial (ca. 120.000–140.000 before present, BP);
LGM-CCSM und LGM-MIROC, letztes glaziales Maximum (ca. 21.000 BP) und
PRE, heutige Bedingungen (ca. 1950–2000).
Dunklere Farben zeigen eine höhere Wahrscheinlichkeit passender Klimabedingungen. Gebiete mit mittlerer, niedriger oder keiner klimatischen Eignung sind grau dargestellt.
Im Gegensatz zu früheren ökologischen Studien, die die Bedeutung der Wasserversorgung für die demographischen Prozesse von T. baccata (z.B. Naturverjüngung oder Geschlechterverhältnis einer Population) hervorheben, zeigt diese Studie keine direkte Wirkung der Niederschlagsmenge auf die genetische Entwicklung, wohl aber einen bedeutenden Einfluss der Temperatur. Es existiert „eine signifikante Verbindung von Geschlechterverhältnis und Temperatur, wobei die westlichen (westlicher Mittelmeerraum und Britische Inseln) und östlichen (Zentral- und Nordeuropa) Populationen deutlich zwei unterschiedliche Gruppen darstellen (…). Das weist auf die Existenz zweier evolutionärer Abstammungslinien hin, die sich an entgegengesetzte Temperaturbedingungen anpassten“.
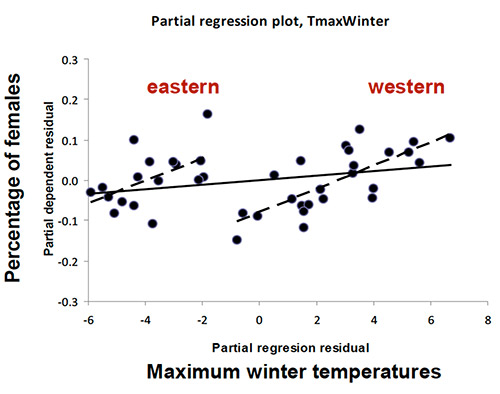
Die Populationen des westlichen Genpools haben ganz ähnliche Geschlechterverhältnisse wie die östlichen, aber bei höheren Temperaturen.
Schlussfolgerung
„Die Herausarbeitung des genauen Beitrags verschiedener Interglaziale (PRE und LIG) auf umweltgetriebene Isolation ist eine Herausforderung, vor allem in Abwesenheit umfassender Fossilienfunde. Auch wenn wir eine temporär variierende Auswahl nicht ausschließen können, gibt es einige Belege dafür, dass sich adaptive Prozesse höchstwahrscheinlich während des letzten Interglazials ereignet haben.“
Die Ergebnisse dieser Arbeit geben eine klare Perspektive auf den klimatischen Einfluss der quatärneren Vereisungen und darauf, wie Selektionsdruck während der Warmzeiten weitere Auswirkungen auf die genetische Entwicklung isolierter Populationen erzeugt haben könnte. „Dies eröffnet neue Untersuchungsmethoden, um die Auswirkung quatärnerer Klimafaktoren auf die heutigen Verbreitungsmuster der genetischen Vielfalt in anderen langlebigen Organismen zu erforschen.“
Literatur
Maria Mayol, Miquel Riba, Santiago C. Gonzalez-Martınez, Francesca Bagnoli, Jacques-Louis de Beaulieu, Elisa Berganzo, Concetta Burgarella, Marta Dubreuil, Diana Krajmerova, Ladislav Paule, Ivana Romsakova, Cristina Vettori, Lucie Vincenot and Giovanni G. Vendramin 2015. Adapting through glacial cycles: insights from a long-lived tree (Taxus baccata). New Phytologist (2015), doi: 10.1111/nph.13496














